


احدث الاخباراخبار مصرالصحهالعالمتكنولوجيارأيعلم النفس سيكولوجيقسم تنمية الموارد البشريةمركز الدراسات المستقبليةمنوعات
لماذا تحتوي الخلايا العصبية على آلاف المشابك (بحث علمي )
نظرية ذاكرة التسلسل في القشرة المخية الحديثة (أساس خوارزمية HTM)
متابعة / أماني عزت
تمثل الخلايا العصبية الهرمية معظم الخلايا العصبية المثيرة في القشرة المخية
الحديثة. يتلقى كل خلية عصبية هرمية مدخلات من آلاف من المشابك المثيرة
المنفصلة إلى فروع من التشعبات. تنقسم التشعبات نفسها إلى مناطق تكامل
قمي وقاعدية وقريبة ، والتي لها خصائص مختلفة. كيف تدمج الخلايا العصبية
الهرمية المدخلات من آلاف نقاط الاشتباك العصبي ، وما هو الدور الذي تلعبه
التشعبات المختلفة في هذا التكامل ، وما نوع سلوك الشبكة الذي يلعبه هذا
في الأنسجة القشرية هو لغز. تم اقتراح سابقًا أن الخصائص غير الخطية
للتشعبات تمكن الخلايا العصبية القشرية من التعرف على أنماط مستقلة
متعددة. في هذا المقال ، قمنا بتوسيع هذه الفكرة بعدة طرق. أولاً ، أظهرنا أنه
حتى في ظل وجود الكثير من الضوضاء وتغيرات الأنماط ، يمكن للخلايا العصبية
التي تحتوي على آلاف المشابك المعزولة في التشعبات النشطة التعرف على
مئات أنماط نشاط الخلية المستقلة. بعد ذلك ، نقترح نموذجًا للخلايا العصبية
تؤدي فيه الأنماط المكتشفة على التشعبات القريبة إلى إمكانات فعلية ، وتحدد
المجال الاستقبالي الكلاسيكي للخلايا العصبية ، وتكون الأنماط المكتشفة على
التشعبات القاعدية والقمية غير مستقطبة قليلاً بواسطة تلعب الخلايا العصبية
دورًا تنبئيًا دون توليد إمكانات فعلية. من خلال هذه الآلية ، يمكن للخلايا العصبية
أن تتنبأ بتنشيطها في مئات البيئات المستقلة. بعد ذلك ، نقترح نموذج شبكة
قائم على الخلايا العصبية يتميز بخاصية تعلم التسلسلات المستندة إلى الوقت.
تعتمد الشبكة على تثبيط موضعي سريع لتنشيط الخلايا العصبية المستقطبة
قليلاً بشكل تفضيلي. من خلال المحاكاة ، نوضح أنه طالما أن الشبكة تستخدم
رمزًا موزعًا للتنشيط الخلوي المتناثر ، يمكن للشبكة أن تتوسع بشكل جيد
وتعمل ضمن نطاقات معلمات عديدة. قارنا خصائص نموذج الشبكة الجديد بالعديد
من نماذج الشبكات العصبية الأخرى لتوضيح القدرات النسبية لكل نموذج.
نستنتج أن الخلايا العصبية الهرمية مع الآلاف من نقاط الاشتباك العصبي
والتشعبات النشطة ومناطق التكامل المتعددة تخلق ذاكرة تسلسل قوية وقوية.
في ضوء الشمولية والتشابه بين الخلايا العصبية المثيرة في القشرة المخية
الحديثة بأكملها وأهمية ذاكرة التسلسل في التفكير والسلوك ، نقترح أن هذا
الشكل من ذاكرة التسلسل قد يكون سمة مشتركة للأنسجة القشرة الحديثة.
نظرة عامة
تحتوي الخلايا العصبية المثيرة في القشرة المخية الحديثة على آلاف من نقاط
الاشتباك العصبي المثيرة. المشبك القريب ، أي المشبك الأقرب إلى جسم
الخلية ، له تأثير كبير نسبيًا على إمكانية توليد الخلية لإمكانات العمل. ومع ذلك ،
فإن معظم نقاط الاشتباك العصبي بعيدة أو بعيدة عن جسم الخلية. تنشيط
المشبك البعيدة الوحيد له تأثير ضئيل على الجسم ، ومن الصعب لسنوات عديدة
تخيل كيف تلعب آلاف المشابك البعيدة دورًا مهمًا في تحديد الاستجابات الخلوية
(Major et al. ، 2013). لقد لوحظ أن الفروع التغصنية هي عناصر معالجة
نشطة. يمكن أن يؤدي تنشيط العديد من المشابك البعيدة في القرب المكاني
والزماني القريب إلى قمم NMDA المتشعبة المحلية وبالتالي إلى إزالة
الاستقطاب الكبير والمستمر للخلايا الجسدية (Antic et al. ، 2010 ؛ Major et
al. ، 2013 ). وقد أدى ذلك ببعض الباحثين إلى اقتراح الفروع المتغصنة
كمعرفات أنماط مستقلة (Poirazi et al. ، 2003 ؛ Polsky et al. ، 2004) ومع
ذلك ، بالمقارنة مع شبكة متعددة الطبقات من الخلايا العصبية بدون التشعبات
النشطة ، فإن الفوائد الوظيفية والنظرية للشبكات العصبية ذات التشعبات
النشطة غير واضحة (Poirazi et al. ، 2003).
ملاحظة: NMDA الجهاز العصبي المركزي المثير للأحماض الأمينية مستقبلات
التأين. يمكن أن يؤدي تنشيط مستقبلات NMDA إلى زيادة نفاذية أغشية الخلايا
العصبية وتوليد إمكانات ما بعد المشبكية المثيرة.
يفتقر إلى النظرية القائلة بأن الخلايا العصبية تحتاج إلى تشعبات نشطة. تقريبًا
جميع الشبكات العصبية الاصطناعية ، مثل تلك المستخدمة في التعلم العميق
(LeCun et al.، 2015) والشبكات العصبية المرتفعة (Maass، 1997) ، هي
باستخدام الخلايا العصبية الاصطناعية مع النماذج التغصنية المبسطة ، قد تفتقر
إلى إمكانية الوظائف الرئيسية للنسيج العصبي البيولوجي. من أجل فهم كيفية
عمل القشرة المخية الحديثة وبناء نظام له نفس مبادئ القشرة المخية الحديثة ،
نحتاج إلى فهم كيفية دمج الخلايا العصبية البيولوجية المدخلات من آلاف
المشابك العصبية وما إذا كانت التشعبات النشطة تلعب دورًا مهمًا. بالطبع ، لا
يمكن فهم الخلايا العصبية بمعزل عن غيرها. لذلك ، نحتاج أيضًا إلى نظرية
تكميلية ، أي كيف تعمل كل شبكة عصبية والتشعبات النشطة معًا لتحقيق هدف
مشترك.
في هذه المقالة ، قدمنا مثل هذه النظرية. أولاً ، أظهرنا كيف تتعرف الخلايا
العصبية الهرمية النموذجية ذات التشعبات النشطة والآلاف من نقاط الاشتباك
العصبي على مئات أنماط نشاط الخلية المستقلة. نظهر ذلك
المتطلب الأساسي: طالما أن النشاط العصبي الإجمالي ضئيل ، حتى لو كان
هناك الكثير من الضوضاء والتنوع ،
الوظيفة: يمكن للخلايا العصبية أيضًا التعرف على هذه الأنماط المستقلة.
نقدم نموذجًا للخلايا العصبية :،
حيث يتم استخدام مدخلات أجزاء مختلفة من الشجرة التغصنية لأغراض مختلفة.
في هذا النموذج ، تؤدي الأنماط المكتشفة على التشعبات القريبة إلى إمكانات
العمل ، والتي تحدد المجال الاستقبالي الكلاسيكي للخلايا العصبية. تلعب
الأنماط التي تتعرف عليها المشابك العصبية البعيدة دورًا تنبئيًا عن طريق إزالة
استقطاب الخلية دون التسبب مباشرة في إمكانات الفعل. من خلال هذه الآلية ،
يمكن للخلايا العصبية أن تتعلم التنبؤ بتنشيطها في مئات السياقات الفريدة. ثم
أوضحنا كيف تتعلم هذه الشبكات العصبية وتتذكر تسلسلات الأنماط. تعتمد
الشبكة على عملية تنافسية تصدر فيها الخلايا العصبية المستقطبة سابقًا
طفرات أسرع من الخلايا العصبية غير المستقطبة. عندما يقترن بالتثبيط المحلي
السريع ، فإن حالة تنشيط الشبكة تكون منحازة نحو توقعها. تشكل دورات
التنشيط التي تؤدي إلى التنبؤ والتفعيل وما إلى ذلك أساس الذاكرة
المتسلسلة.
نصف مجموعة من قواعد التعلم والتفعيل المطلوبة للخلايا العصبية ذات
التشعبات النشطة. لا يمكن لشبكات الخلايا العصبية الخطية أو غير الخطية
القياسية ذات الهياكل التغصنية المبسطة تنفيذ قواعد التنشيط والتعلم هذه
بسهولة. للقيام بذلك ، يجب إدخال بعض الميزات المعقدة وغير المحتملة من
الناحية البيولوجية. لذلك ، اخترنا استخدام نماذج الخلايا العصبية التي تتضمن
التشعبات النشطة ومناطق التكامل للتشعبات القريبة والقاعدية والقمية ،
ونعتقد أنها تتطابق بشكل وثيق مع تشريح الخلايا العصبية وعلم وظائف الأعضاء
المعروفين.
من خلال المحاكاة ، نوضح أن شبكة الذاكرة المتسلسلة تعرض العديد من
الخصائص المتوقعة ، مثل التعلم عبر الإنترنت والتنبؤات المتعددة المتزامنة
والمتانة. تتوافق النظرية الكلية مع كمية كبيرة من الأدلة التجريبية. نحدد بعض
التنبؤات البيولوجية التفصيلية التي يمكن استخدامها لمزيد من اختبار النظرية.
النتائج
تتعرف الخلايا العصبية بشكل موثوق على أنماط متفرقة متعددة ، وعادة ما يُنظر
إلى الخلايا العصبية على أنها تحسب مجموعًا مرجحًا واحدًا لجميع نقاط
الاشتباك العصبي. هذا المفهوم ، الذي يطلق عليه أحيانًا “النقطة العصبية” ،
يشكل أساس جميع الشبكات العصبية الاصطناعية تقريبًا (الشكل 1 أ).
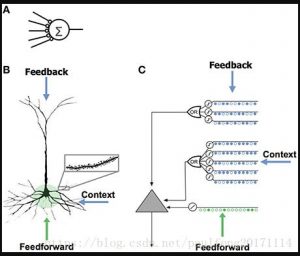
الشكل 1. مقارنة بين نماذج الخلايا العصبية. (أ) نموذج الخلايا العصبية
المستخدم في معظم الشبكات العصبية الاصطناعية يحتوي على عدد قليل من
نقاط الاشتباك العصبي وعدم وجود تشعبات. (ب) الخلايا العصبية الهرمية
القشرية الحديثة لديها آلاف من نقاط الاشتباك العصبي المثيرة الموجودة على
التشعبات (أقحم). سيؤدي التنشيط المشترك لمجموعة من نقاط الاشتباك
العصبي على شظايا شجرية إلى حدوث قمم NMDA وإزالة الاستقطاب
الجسدي. تحتوي الوحدة على ثلاثة مصادر إدخال. المدخلات التغذوية (الموضحة
باللون الأخضر) أشكال التشابك العصبي بالقرب من الخلايا الجسدية ، مما يؤدي
مباشرة إلى إمكانات الفعل. إن طفرات NMDA التي يتم إنتاجها في التشعبات
القاعدية والقمية البعيدة تزيل استقطاب الخلايا الجسدية ، ولكنها عادة لا تكون
كافية لتوليد إمكانات فعل جسدية. (ج) تحاكي الخلايا العصبية لنموذج HTM
التشعبات ومسامير NMDA ، والتي تحتوي على سلسلة من أجهزة الكشف
المتزامنة ، كل منها مع مجموعة من نقاط الاشتباك العصبي (يتم عرض عدد
قليل فقط من كل مشابك).
تقترح التشعبات النشطة وجهات نظر مختلفة للخلايا العصبية ، حيث تتعرف
الخلايا العصبية على العديد من الأنماط الفريدة المستقلة (Poirazi et al.، 2003؛
Polsky et al.، 2004؛ Larkum and Nevian، 2008). تُظهر النتائج التجريبية أن
التنشيط المتزامن لـ 8-20 من نقاط الاشتباك العصبي في القرب المكاني
القريب على التشعبات سوف يتحد بطريقة غير خطية ويسبب طفرات شجرية
NMDA (لاركوم وآخرون ، 1999 ؛ شيلر وآخرون ، 2000 ؛ شيلر وآخرون). شيلر ،
2001 ؛ ميجور وآخرون ، 2013). لذلك ، تعمل مجموعة صغيرة من المشابك
المجاورة ككاشفات للأنماط. يمكن ملاحظة أن الآلاف من نقاط الاشتباك العصبي
على التشعبات الخلوية تعمل كمجموعة من أجهزة الكشف عن الأنماط
المستقلة. سيؤدي اكتشاف أي من هذه الأنماط إلى ظهور قمم NMDA وإزالة
الاستقطاب الجسدي اللاحقة.
يبدو أن 8-20 من نقاط الاشتباك العصبي لا يمكنها تحديد أنماط النشاط بشكل
موثوق في عدد كبير من مجموعات الخلايا. ومع ذلك ، إذا كان النمط المراد
التعرف عليه ضئيلًا ، فيمكن إجراء التعرف القوي ؛ أي ، بالنسبة إلى السكان ،
فإن عددًا قليلاً من الخلايا العصبية النشطة (Olshausen and Field ، 2004).
على سبيل المثال ، ضع في اعتبارك مجموعة مكونة من 200 ألف خلية ، حيث
تكون 1٪ (2000) من الخلايا نشطة في أي وقت. نريد أن تكتشف الخلايا
العصبية متى تظهر أنماط معينة في 200 ألف خلية. إذا كان جزء من التشعبات
في الخلايا العصبية يشكل فقط نقاط الاشتباك العصبي الجديدة لـ 10 من أصل
2000 خلية نشطة ، وكان الحد الأدنى لتوليد طفرات NMDA هو 10 ، عندما
تتلقى جميع نقاط الاشتباك العصبي العشر التنشيط في نفس الوقت ،
ستكتشف التشعبات النمط المستهدف . لاحظ أن التشعبات قد تكتشف بشكل
غير صحيح العديد من الأنماط الأخرى التي تشترك في نفس الوحدات العشر
النشطة. ومع ذلك ، إذا كانت الأنماط متفرقة ، فهناك احتمال ضئيل بأن تصبح 10
نقاط تشابك عصبية نشطة لأنماط عشوائية مختلفة. في هذا المثال لديها
فقط 9.8×10−21 9.8 × 10 − 21 。
يمكن حساب احتمال المطابقة غير الصحيحة تمامًا كما يلي. افترض n n يمثل
حجم مجموعة الخلايا في نقطة زمنية معينة و α α عدد الخلايا النشطة في هذا
المجتمع ، للوضع المتناثر α≪n α ≪ n . افترض s s هو عدد نقاط الاشتباك العصبي في
الجزء الشجيري ، θ θ هي عتبة ذروة NMDA. نقول لو على الأقل θ θ يصبح المشبك
نشطًا ، ثم وضع التعرف على الجزء ، أي s s متشابك على الأقل θ θ تطابق مع
الخلية النشطة حاليا.
بافتراض التوزيع العشوائي للنمط ، يتم تحديد الاحتمال الدقيق للتطابق الخاطئ
من خلال:
limsb=θ(sb)×(n−sa−b)(na)
lim b = θ s ( s b ) × ( n − s a − b ) ( n a )
(1)
المقام فقط في n n العدد الإجمالي للأنماط الممكنة للخلايا القابلة للحياة في
إجمالي عدد الخلايا. ترتبط الحوسبة الجزيئية بقطعة شجيرية θ θ او اكثر s s عدد
نقاط الاشتباك العصبي. يمكن العثور على وصف أكثر تفصيلاً لهذه المعادلة في
Ahmad and Hawkins (2016).
توضح هذه المعادلة أن التقسيم الشجيري غير الخطي يمكنه تصنيف الأنماط
بقوة عن طريق أخذ عينات فرعية (فقط عدد صغير من الخلايا في النمط يتم
تصنيفها من نقاط الاشتباك العصبي). يسرد الجدول A في نص S1 احتمالية
الخطأ التمثيلية المحسوبة وفقًا للمعادلة (1).

من خلال تكوين المزيد من نقاط الاشتباك العصبي مما هو مطلوب لإنتاج طفرات
NMDA ، يصبح التعرف قويًا على الضوضاء والتغييرات. على سبيل المثال ، إذا
كانت عتبة الذروة التغصنية NMDA هي 10 ، لكنها تشكل 20 نقطة تشابك للنمط
الذي تريد التعرف عليه ، وهو ضعف العدد المطلوب ، فإنه يسمح للتشعبات
بالتعرف على النمط المستهدف حتى إذا تم تغيير 50٪ من الخلايا أو غير نشط.
تزيد المشابك الإضافية أيضًا من احتمال حدوث أخطاء إيجابية خاطئة. على الرغم
من زيادة احتمال الخطأ ، توضح المعادلة (1) أنه لا يزال صغيراً عندما يكون النمط
متفرقًا. في المثال أعلاه ، مضاعفة عدد نقاط الاشتباك العصبي وبالتالي إدخال
هامش ضوضاء بنسبة 50٪ يزيد من فرصة الخطأ فقط 1.6×10−18 1.6 × 10 − 18 . يسرد
الجدول B في نص S1 معدل الخطأ التمثيلي عندما يتجاوز عدد نقاط الاشتباك
العصبي العتبة.
يجب وضع نقاط الاشتباك العصبي التي تتعرف على نمط معين في مكان واحد
على مقاطع شجيرية. إذا كانت تبعد 40 ميكرون عن بعضها البعض ، فإن ما لا
يقل عن 8 مشابك كهربائية كافية لإنتاج طفرات NMDA (Major et al. ، 2008).
إذا تم توزيع نقاط الاشتباك العصبي على طول العقد التغصنية ، فستكون هناك
حاجة إلى ما يصل إلى 20 نقطة تشابك عصبية (Major et al. ، 2013). يمكن أن
تحتوي المقاطع الشجيرية على مئات المشابك ؛ لذلك ، يمكن لكل جزء اكتشاف
أنماط متعددة. إذا تم مزج المشابك التي تتعرف على الأنماط المختلفة معًا على
شظايا شجيرية ، يتم تقديم احتمالات خطأ إضافية عن طريق التنشيط المشترك
لنقاط الاشتباك العصبي من أنماط مختلفة. يعتمد احتمال هذا النوع من الخطأ
على عدد مجموعات نقاط الاشتباك العصبي التي تشترك في المقاطع
المتغصنة وتناثر النمط المراد التعرف عليه. بالنسبة لمجموعة كبيرة من القيم ، لا
يزال احتمال حدوث مثل هذه الأخطاء منخفضًا (الجدول C في نص S1). لذلك ،
فإن وضع نقاط الاشتباك العصبي التي تتعرف على أنماط معينة يكون دقيقًا إلى
حد ما (يجب أن تكون على نفس المقطع الشجيري وفي حدود 40 ميكرومتر من
بعضها البعض بشكل مثالي) ، ولكنها أيضًا غير دقيقة إلى حد ما (لا تختلط مع
المشابك الأخرى) قد يسبب أخطاء).
إذا افترضنا أن 20 نقطة تشابك يتم توزيعها بالتساوي للتعرف على كل نمط ، وأن
الخلايا العصبية بها 6000 نقطة تشابك عصبية ، فإن الخلية ستتمتع بالقدرة على
التعرف على حوالي 300 نمط مختلف. هذا تقدير تقريبي ، لكن من الواضح أن
الخلايا العصبية ذات التشعبات النشطة يمكنها أن تتعلم بشكل موثوق التعرف
على مئات الأنماط في مجموعات الخلايا الكبيرة. سيؤدي التعرف على أي من
هذه الأنماط إلى إزالة استقطاب الخلية. نظرًا لأن جميع الخلايا العصبية المثيرة
في القشرة المخية الحديثة تحتوي على آلاف من نقاط الاشتباك العصبي ،
وبقدر ما نعلم ، فكلها تحتوي على تشعبات نشطة ، فإن كل خلية عصبية مثيرة
للقشرة المخية الحديثة يمكنها التعرف على مئات الأنشطة العصبية الوضع.
في القسم التالي ، نقترح أن معظم الأنماط التي تتعرف عليها الخلايا العصبية لا
تؤدي مباشرة إلى إمكانات العمل ، ولكنها تلعب دورًا في كيفية توقع الشبكة
العصبية للتسلسلات وتعلمها.
ثلاثة مصادر للمدخلات المشبكية من الخلايا العصبية القشرية
تتلقى الخلايا العصبية مدخلات مثيرة من مصادر مختلفة ، والتي يتم فصلها في
أجزاء مختلفة من الشجرة التغصنية. يوضح الشكل 1 ب الخلايا الهرمية
النموذجية ، وهي الخلايا العصبية الأكثر شيوعًا في القشرة المخية الحديثة.
نقسم مدخلات الخلية إلى ثلاث مناطق. تتلقى المنطقة القريبة النهاية مدخلات
تغذية. تتلقى المنطقة القاعدية مدخلات سياقية ، بشكل أساسي من الخلايا
المجاورة في نفس المنطقة القشرية (Yoshimura et al. ، 2000 ؛ Petreanu et
al. ، 2009 ؛ Rah et al. ، 2013). تتلقى المنطقة العلوية مدخلات التغذية
الراجعة (Spruston ، 2008). (ثاني أكثر الخلايا العصبية الاستثارة شيوعًا في
القشرة المخية الحديثة هي الخلايا النجمية الشوكية ؛ نعتقد أنها تعتبر مشابهة
للخلايا الهرمية مطروحًا منها التشعبات القمية.) نقترح الخلايا العصبية (القريبة
والقاعدية والقمية) ) من مناطق التكامل الثلاثة متشابك. يخدم الأغراض التالية.
يحدد المشبك الداني المجال الاستقبالي الكلاسيكي للخلية
المشابك (عادةً بضع مئات) على التشعبات القريبة لها تأثير كبير نسبيًا في
الخلايا الجسدية ، لذا فهي مناسبة جدًا لتحديد استجابة المجال الاستقبالي
الأساسية للخلايا العصبية (Spruston ، 2008). إذا كان التنشيط المتزامن
للمجموعة الفرعية من المشبك القريب كافياً لتوليد إمكانية فعل جسدية ، وإذا
كان إدخال المشبك القريب نشطًا بشكل ضئيل ، فإن المشبك القريب سيتعرف
على أنماط تغذية فريدة متعددة بنفس الطريقة كما نوقش سابقًا. لذلك ، يمكن
اعتبار مجال استقبال التغذية الأمامي للخلية مزيجًا من وضع التغذية إلى الأمام.
التحولات في تسلسل تعلم المشبك الأساسي
نقترح أن تتعرف التشعبات القاعدية للخلايا العصبية على نمط نشاط الخلية قبل
أن تنطلق الخلايا العصبية ، بحيث تتعلم التشعبات القاعدية وتخزن الانتقال بين
أنماط النشاط. عندما يتم التعرف على نمط على التشعبات القاعدية ، فإنه ينتج
طفرات NMDA. نظرًا لأن إزالة الاستقطاب الناجم عن ارتفاع NMDA يضعف
السعة عندما تصل إلى الخلية الجسدية ، فعندما تتعرف التشعبات القاعدية على
نمط ما ، فإنها ستزيل استقطاب الخلية الجسدية ولكن ليس كافيًا لتوليد جهد
فعل جسدي (Antic et al. ، 2010 ؛ ميجور وآخرون ، 2013). نقترح أن هذا الحد
الأدنى من الاستقطاب هو حالة مهمة للخلية. إنه يمثل تنبؤًا بأن الخلايا ستصبح
نشطة قريبًا وتلعب دورًا مهمًا في سلوك الشبكة. إذا تم تلقي مدخلات تغذية
كافية لاحقًا ، فسوف ترسل خلية خالية من الاستقطاب قليلاً في وقت أبكر من
غير ذلك. من خلال إطلاقه في وقت مبكر ، فإنه يثبط الخلايا المجاورة ، مما يخلق
أنماط نشاط متناثرة للغاية للمدخلات المتوقعة بشكل صحيح. سنشرح هذه
الآلية بشكل كامل في قسم لاحق.
المشابك القمية تثير توقعات من أعلى إلى أسفل
تنتج التشعبات القمية للخلايا العصبية أيضًا قمم NMDA عند التعرف على الأنماط
(Cichon and Gan ، 2015). لا يؤثر ارتفاع NMDA العلوي بشكل مباشر على
الخلايا الجسدية. بدلاً من ذلك ، يمكن أن يتسبب في حدوث طفرات Ca2 + في
التشعبات القمية (Golding et al. ، 1999 ؛ Larkum et al. ، 2009). سيؤدي
ارتفاع Ca2 + القمي المفرد إلى إزالة استقطاب الخلايا الجسدية ، ولكنه عادة لا
يكفي لتوليد إمكانات فعل جسدية (Antic et al. ، 2010). يعتبر التفاعل بين
مسامير Ca2 + القمية ، وقمم NMDA القاعدية وإمكانات الفعل الجسدي مجالًا
للبحث المستمر (Larkum ، 2013) ، ولكن يمكننا القول أنه في ظل العديد من
الظروف ، فإن النمط المعترف به على التشعبات القمية سيجعل الخلايا لذلك ،
فإن إزالة الاستقطاب له تأثير مشابه لنمط التعرف على التشعبات الركيزة. نقترح
أن يتم استخدام إزالة الاستقطاب الناتجة عن التشعبات القمية لإنشاء توقعات
من أعلى إلى أسفل ، والتي يمكن اعتبارها شكلاً آخر من أشكال التنبؤ.
نموذج عصبون HTM
يوضح الشكل 1C النموذج المجرد للخلايا العصبية الهرمية التي استخدمناها في
محاكاة البرامج. نقوم بنمذجة التشعبات في الخلية كمجموعة من أجهزة كشف
الصدفة العتبة ؛ لكل منها المشبك الخاص بها. إذا تجاوز عدد المشابك النشطة
على كاشف التغصن / الصدفة العتبة ، تكتشف الخلية نمطًا. تنقسم كاشفات
المصادفة إلى ثلاث مجموعات ، تتوافق مع التشعبات القريبة والقاعدية والقمية
للخلايا الهرمية. نسمي هذا النموذج العصبوني “عصبون HTM” لتمييزه عن
الخلايا العصبية البيولوجية والخلايا العصبية النقطية. HTM هو اختصار للذاكرة
الزمنية الهرمية ويستخدم لوصف نموذجنا القشري الجديد (هوكينز وآخرون ،
2011). تحتوي الخلايا العصبية HTM المستخدمة في عمليات المحاكاة هنا على
128 كاشفًا للتشعبات / المصادفة ، ولكل تغصن ما يصل إلى 40 نقطة تشابك
عصبية. من أجل الوضوح ، لا يُظهر الشكل 1C سوى عدد قليل من التشعبات
والمشابك.
تسلسل تعلم الشبكة العصبية
نظرًا لأن جميع الأنسجة في القشرة المخية الحديثة تتكون من خلايا عصبية ذات
تشعبات نشطة وآلاف من نقاط الاشتباك العصبي ، فهذا يدل على وجود جميع
مبادئ شبكة القشرة المخية الحديثة. هذا يقودنا إلى السؤال ، ما هي خصائص
الشبكة الأساسية لدرجة أنها جزء ضروري من التفكير الحسي والتنبؤ واللغة
والتخطيط الحركي؟
نقترح أن العملية الأساسية لجميع أنسجة القشرة المخية الحديثة هي تعلم
واسترجاع تسلسلات الأنماط (Hawkins and Blakeslee ، 2004) ، وهو ما
يسميه كارل لاشلي “أهم مشكلة فيزيولوجيا الدماغ وأكثرها إغفالًا” (لاشلي ،
1951). وبشكل أكثر تحديدًا ، نقترح أن تنفذ كل طبقة خلية في القشرة المخية
الحديثة متغيرًا من خوارزمية ذاكرة التسلسل الشائعة. نوصي بأن تستخدم
طبقة الخلية ذاكرة التسلسل لأغراض مختلفة ، وهذا هو سبب تغيير طبقة
الخلية في التفاصيل ، مثل الحجم والاتصال. في هذه المقالة ، نشرح ما نعتقد
أنه خوارزمية ذاكرة التسلسل الأساسية ، دون شرح تغييراتها بالتفصيل.
نبدأ استكشاف ذاكرة التسلسل من خلال سرد العديد من السمات التي تتطلبها
الشبكة لنمذجة القشرة المخية الحديثة.
(1) التعلم عبر الإنترنت
يجب أن يكون التعلم مستمرًا. إذا تغيرت إحصاءات العالم ، يجب أن تتكيف الشبكة
تدريجياً وباستمرار مع كل مدخلات جديدة.
(2) التنبؤ عالي المستوى
يتطلب استخدام التسلسلات المعقدة لعمل تنبؤات صحيحة القدرة على دمج
المعلومات السياقية السابقة. تحتاج الشبكة إلى تحديد مقدار الوقت المطلوب
بشكل ديناميكي لتحقيق أفضل توقع. يشير مصطلح “ترتيب أعلى” إلى
“سلسلة ماركوف ذات رتبة أعلى” بهذه الخاصية.
(3) تنبؤات متعددة في وقت واحد
عادةً ما تحتوي تدفقات البيانات الطبيعية على تسلسلات متداخلة ومتفرعة.
لذلك ، تحتاج ذاكرة التسلسل إلى إجراء تنبؤات متعددة في نفس الوقت.
(4) قواعد التعلم المحلية
يجب أن تستخدم ذاكرة التسلسل قواعد التعلم المحلية فقط لكل خلية عصبية.
يجب أن تكون القواعد محلية في المكان والزمان ، دون الحاجة إلى وظيفة
موضوعية عالمية.
(5) المتانة
يجب أن تُظهر الذاكرة متانة لمستويات عالية من الضوضاء وفقدان الخلايا
العصبية والتغيرات الطبيعية في الإدخال. يجب أن يكون تدهور الأداء في ظل
هذه الظروف تدريجيًا.
يجب أن تحدث كل هذه السمات في وقت واحد في سياق بيانات التدفق
المستمر.
أعمدة صغيرة وخلايا عصبية: تمثيلان
تتطلب ذاكرة التسلسل عالية المستوى تمثيلين متزامنين. أحدهما يمثل
المدخلات العارضة للشبكة ، والآخر يمثل المدخلات المغذية في سياق زمني
محدد. لتوضيح هذا المطلب ، ضع في اعتبارك تسلسلين مجردين هما “ABCD” و
“XBCY” ، حيث يمثل كل حرف نمط التنشيط المتناثر في مجتمع الخلايا العصبية.
بمجرد معرفة هذه التسلسلات ، يجب أن تتنبأ الشبكة بـ “D” عند تقديم
التسلسل “ABC” ، ويجب أن تتنبأ بـ “Y” عند تقديم التسلسل “XBC”. لذلك ،
يجب أن يكون التمثيل الداخلي أثناء “BC” التالي حالتين مختلفتين ؛ وإلا ، لا
يمكن إجراء التنبؤ الصحيح بعد تقديم “C”.
يوضح الشكل 2 كيف نقترح أن يتم تمثيل هذين التمثيلين في طبقة خلية الخلايا
العصبية القشرية. تمثل اللوحة في الشكل 2 شريحة من خلال طبقة خلية
واحدة في القشرة المخية الحديثة (الشكل 2 أ). من أجل الوضوح ، تم تبسيط
اللوحة بشكل كبير. يوضح الشكل 2 ب كيف تمثل الشبكة تسلسلي الإدخال قبل
تعلم التسلسل. يوضح الشكل 2 ج كيف تمثل الشبكة نفس المدخلات بعد
تسلسل التعلم. يتم تحويل كل مدخلات تلقائية للشبكة إلى مجموعة من أعمدة
النشاط الصغيرة المتفرقة. (تمتد الأعمدة الصغيرة في القشرة المخية الحديثة
على طبقات خلايا متعددة. هنا نشير فقط إلى الخلايا الموجودة في الأعمدة في
طبقة خلية واحدة). تشترك جميع الخلايا العصبية في الأعمدة الصغيرة في نفس
المجال الاستقبالي الأمامي. في حالة وصول إدخال عرضي ، ستتعرف جميع
الخلايا الموجودة في العمود المصغر المحدد على وضع الإدخال وتصبح نشطة.
ومع ذلك ، في سياق التسلسل الذي تم تعلمه سابقًا ، سيتم إزالة الاستقطاب
خلية واحدة أو أكثر في العمود الصغير. ستولد الخلايا غير المستقطبة أولاً
إمكانات فعلية ، مما يثبط الخلايا الأخرى المجاورة. لذلك ، سينتج عن الإدخال
المتوقع نمطًا متناثرًا جدًا من تنشيط الخلية ، وهو فريد لعنصر معين ، في موقع
معين ، وفي تسلسل محدد.
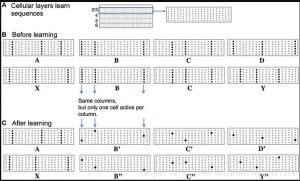
الشكل 2. يظهر التسلسل في طبقة الخلية القشرية. (أ) تنقسم القشرة المخية
الحديثة إلى طبقات خلوية. تعرض اللوحة في هذا الشكل جزءًا من طبقة خلية
عامة. من أجل الوضوح ، تعرض اللوحة 21 شريطًا صغيرًا فقط ، كل منها يحتوي
على 6 خلايا. (ب) لم يتم التعرف على تسلسل الإدخال ABCD و XBCY بعد.
يستدعي كل عنصر تسلسل مجموعة من الأعمدة الصغيرة المتفرقة ، وفي هذا
الشكل يوجد ثلاثة فقط. إذا كان الإدخال غير متوقع ، تصبح جميع الخلايا في
العمود الصغير نشطة ، وهذا هو الحال قبل تسلسل التعلم. (ج) بعد تعلم
التسلسلين ، يستدعي الإدخال نفس الأعمدة المصغرة ، لكن وحدة واحدة فقط
في كل عمود صالحة ، تم وضع علامة B ‘، B “، C’ ، C” ، D ‘و Y “. لأن C’ و C
“فريدة ، لذا يمكنهم استدعاء التنبؤ الصحيح عالي الترتيب لـ Y أو D.
المشابك القاعدية هي أساس ذاكرة التسلسل
في هذه النظرية ، تستخدم الخلايا المشابك القاعدية لتعلم التبديل بين أنماط
الإدخال. مع كل إدخال تغذية جديد ، تصبح بعض الخلايا نشطة من خلال نقاط
الاشتباك العصبي القريبة. تستخدم الخلايا الأخرى نقاط الاشتباك العصبي
الأساسية الخاصة بها لتتعلم التعرف على هذا النمط من النشاط ، وعندما ترى
النمط مرة أخرى ، فإنها تصبح خالية من الاستقطاب ، وتتنبأ بتنشيطها المغذي
في الإدخال التالي. ينشط الإدخال الأمامي الخلية ، ويُنتج الإدخال الأساسي
تنبؤات. طالما أن الإدخال التالي يطابق التنبؤ الحالي ، سيستمر التسلسل ، كما
هو موضح في الشكل 3. يوضح الشكل 3 أ الخلايا النشطة والخلايا المتوقعة ،
بينما تتبع الشبكة التسلسل الذي تم تعلمه مسبقًا.

الشكل 3. تتنبأ الوصلات الأساسية بالخلايا العصبية القريبة بالإدخال التالي. (أ)
باستخدام تسلسل من الشكل 2 ، يظهر الخلايا النشطة (السوداء) والخلايا غير
المستقطبة / المتوقعة (باللون الأحمر). تُظهر اللوحة الأولى الإدخال العرضي A
، مما يؤدي إلى توقع الإدخال التالي B ‘(اللوحة الثانية). إذا تطابق الإدخال
اللاحق مع التنبؤ ، فإن الوحدة المزيلة للاستقطاب هي فقط التي ستصبح
نشطة (اللوحة الثالثة) ، مما يؤدي إلى تنبؤ جديد (اللوحة الرابعة). تظهر إحدى
الوصلات المشبكية الجانبية التي تستخدمها الخلية المتوقعة في الصورة
الموجودة في أقصى اليمين. في شبكة حقيقية ، سيكون لكل خلية متوقعة 15
اتصالاً أو أكثر بمجموعة فرعية من عدد كبير من الخلايا النشطة. (ب) قدم
التسلسل الفرعي الغامض “BC” (وهو جزء من ABCD و XBCY) للشبكة. تُظهر
اللوحة الأولى المدخلات غير المتوقعة لـ B ، مما يؤدي إلى التنبؤ بـ C’و C “. تُظهر
اللوحة الثالثة النظام بعد الإدخال C. تصبح مجموعتا الوحدات المتوقعة نشطة ،
مما يؤدي إلى التنبؤ بـ D و Y (المجموعة الرابعة لوحة) في تدفقات البيانات
المعقدة ، عادة ما يكون هناك العديد من التنبؤات المتزامنة.
عادة ، تقوم الشبكة بعمل تنبؤات متعددة في وقت واحد. على سبيل المثال ،
لنفترض أنه بعد تعلم التسلسل “ABCD” و “XBCY” ، فإننا نعرض النظام
للتسلسل الفرعي الضبابي “BC”. في هذه الحالة ، نريد أن يتوقع النظام “D” و
“Y” في نفس الوقت. يوضح الشكل 3 ب كيف تقوم الشبكة بعمل تنبؤات متعددة
عندما يكون الإدخال غير واضح. يمكن حساب عدد التنبؤات المتزامنة التي يمكن
إجراؤها باحتمالية خطأ منخفضة مرة أخرى بواسطة المعادلة (1). نظرًا لأن
التنبؤات غالبًا ما تكون متفرقة جدًا ، يمكن للشبكة التنبؤ بالعشرات من الأوضاع
في نفس الوقت دون ارتباك. إذا تطابق الإدخال مع أي توقع ، فسيؤدي ذلك إلى
تمثيل صحيح متناثر للغاية. إذا كان الإدخال لا يتطابق مع أي تنبؤات ، فستصبح
جميع الخلايا في العمود نشطة ، مما يشير إلى إدخال عرضي.
على الرغم من أن كل خلية في العمود المصغر تشترك في نفس الاستجابة
الأمامية ، فإن نقاط الاشتباك العصبي الخاصة بها تتعرف على أنماط مختلفة.
لذلك ، ستستجيب الوحدات الموجودة في العمود المصغر بشكل فريد في
سياقات وقت التعلم المختلفة ، وعندما يكون الإدخال متوقعًا ، سيكون مستوى
النشاط الإجمالي متناثرًا. وقد لوحظت هاتان الخاصيتان (Vinje and Gallant،
2002؛ Yen et al.، 2007؛ Martin and Schröder، 2013).
بالنسبة لخلية في اللوحة الأخيرة من الشكل 3 أ ، نعرض ثلاثة اتصالات للخلية
المستخدمة للتنبؤ. في خلية عصبية حقيقية ، في محاكاتنا ، ستشكل الخلية
من 15 إلى 40 اتصالًا مع مجموعة فرعية من مجموعة الخلايا النشطة الأكبر.
يخلق المشبك العلوي توقعًا من أعلى إلى أسفل
عادة ما تشكل محاور التغذية الراجعة بين مناطق القشرة الحديثة نقاط الاشتباك
العصبي (في الطبقة 1) والتشعبات القمية للخلايا العصبية الهرمية ، مع وجود
أجسام خلوية في الطبقات 2 و 3 و 5. . لطالما تم التكهن بأن هذه الاتصالات
المرتدة تحقق شكلاً من أشكال التوقعات أو التحيز (Lamme et al. ، 1998).
يقترح نموذجنا العصبي آلية التوقعات التنازلية في القشرة المخية الحديثة.
يوضح الشكل 4 كيف يمكن لنموذج التغذية المرتدة المستقر للتشعبات القمية أن
يتنبأ بعناصر متعددة في التسلسل في وقت واحد. عند وصول مدخلات تغذية
جديدة ، سيتم تفسيرها كجزء من تسلسل التنبؤ. التعليقات تحيز المدخلات نحو
تفسير محدد. أيضًا ، نظرًا لأن الأنماط متفرقة ، يمكن التنبؤ بالعديد من الأنماط
في نفس الوقت.

الشكل 4. التعليقات على التشعبات القمية تتنبأ بالتسلسل بأكمله. يستخدم
هذا الرقم نفس الشبكة والتمثيل كما في الشكل 2. المنطقة المسماة
“التشعبات القمية” تتوافق مع الطبقة 1 في القشرة المخية الحديثة ؛ وتنتهي
التشعبات القمية (غير الموضحة) من جميع الخلايا هنا. في هذا الشكل ، تم
وضع الافتراضات التالية. لقد تعلمت الشبكة مسبقًا التسلسل ABCD ، كما هو
موضح في الشكل 2. أثناء تسلسل التعلم ، يتم تقديم نمط ردود الفعل المستمر
إلى التشعبات القمية ، والخلايا المشاركة في تسلسل شكل B’C’D ‘المشابك
في قمتها. تتعرف التشعبات على أنماط التغذية الراجعة المستمرة. بعد التعرف
على اتصال التغذية الراجعة ، يتم تقديم نمط التغذية الراجعة إلى التشعبات
القمية بينما يتم التعرف عليه من قبل جميع الخلايا في التسلسل الذي سيتم
تنشيطه بالتتابع. تظهر هذه الخلايا باللون الأحمر وتصبح خالية من الاستقطاب
(الجزء الأيسر). عندما يصل إدخال تغذية جديد ، سينتج عنه تمثيل متناثر (اللوحة
الوسطى) متعلق بالتسلسل المتوقع. إذا كان لا يمكن تفسير وضع التغذية
الأمامية كجزء من التسلسل المتوقع (اللوحة اليمنى) ، تصبح جميع الخلايا في
العمود المحدد نشطة ، مما يشير إلى وجود خلل. وبهذه الطريقة ، فإن التعليقات
العليا تحيز الشبكة لتفسير أي إدخال كجزء من التسلسل المتوقع ويكتشف ما
إذا كان الإدخال لا يتطابق مع أي عنصر في التسلسل المتوقع.
لذلك ، هناك نوعان من التنبؤات يحدثان في نفس الوقت. يتنبأ الاتصال الجانبي
بالتغصنات القاعدية بالإدخال التالي ، ويتنبأ الاتصال من أعلى لأسفل بالتغصنات
القمية بالعناصر المتسلسلة المتعددة في نفس الوقت. يعد التفاعل
الفسيولوجي بين التشعبات القمية والقاعدية مجالًا نشطًا للبحث (Larkum ،
2013) وقد يؤدي إلى تفسيرات أكثر دقة لدورها في الاستدلال والتنبؤ. ومع ذلك
، نقترح أن الآلية الموضحة في الشكل 2-4 قد تستمر في لعب دور في التفسير
النهائي.
قواعد التعلم متشابك
يتطلب نموذج الخلايا العصبية الخاص بنا تغييرين في قواعد التعلم التي تعلمتها
معظم النماذج العصبية. أولاً ، يتم التعلم عن طريق نمو وإزالة المشابك من تجمع
المشابك “المحتملة” (Chklovskii et al. ، 2004). ثانيًا ، يحدث التعلم والتغيرات
التشابكية Hebbian على مستوى الأجزاء التغصنية ، وليس على مستوى الخلايا
العصبية بأكملها (Stuart and Häusser ، 2001).
المشبك العصبي المحتمل
لكي تتعرف الخلية العصبية على أنماط النشاط ، فإنها تتطلب مجموعة من نقاط
الاشتباك العصبي المشتركة (عادة 15-20) متصلة بمجموعة فرعية من الخلايا
النشطة في النمط المراد التعرف عليه. يتم تحقيق تعلم التعرف على الأنماط
الجديدة من خلال تكوين مجموعة من نقاط الاشتباك العصبي الجديدة جنبًا إلى
جنب مع مقاطع شجيرية.
يوضح الشكل 5 كيف نقوم بمحاكاة تشكيل نقاط الاشتباك العصبي الجديدة في
الخلايا العصبية HTM. لكل مقطع شجيري ، نحافظ على مجموعة من المشابك
“المحتملة” بين المقطع التغصني والخلايا الأخرى في الشبكة ، وقد تشكل هذه
المشابك نقاط الاشتباك العصبي مع هذا المقطع (Chklovskii et al. عدد نقاط
الاشتباك العصبي المحتملة أكبر من عدد المشابك الفعلية. نقوم بتعيين قيمة
عددية تسمى “الثبات” لكل مشابك عصبية محتملة ، والتي تشير إلى مرحلة
نمو المشبك. تشير قيم الثبات القريبة من الصفر إلى المحاور والتشعبات ، والتي
قد تشكل نقاط الاشتباك العصبي ولكنها لم تبدأ بعد في نمو نقاط الاشتباك
العصبي. تشير قيمة الثبات 1.0 إلى المحاور والتشعبات ذات المشابك الكبيرة
المتكونة بالكامل.
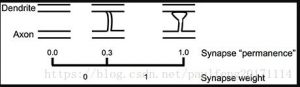
الشكل 5. تعلم عن طريق إضافة نقاط الاشتباك العصبي الجديدة. يتم نمذجة
التعلم في الخلايا العصبية HTM من خلال نمو نقاط الاشتباك العصبي الجديدة
من مجموعة من نقاط الاشتباك العصبي المحتملة. يتم تعيين قيمة “ثبات” لكل
نقطة تشابك محتملة وتمثل نمو المشبك. تعلم عن طريق زيادة أو إنقاص قيمة
المثابرة. إذا كانت الثبات أعلى من العتبة ، فإن وزن المشبك هو قيمة ثنائية يتم
ضبطها على 1.
يستخدم قواعد تشبه Hebbian لزيادة قيمة الاستمرارية وإنقاصها. إذا تجاوزت
قيمة الثبات عتبة ، مثل 0.3 ، فإن وزن المشبك يكون 1 ، وإذا كانت قيمة الثبات
تساوي أو تقل عن العتبة ، فإن وزن المشبك يكون صفرًا. تشير العتبة إلى إنشاء
المشبك ، على الرغم من أنه يمكن للمرء أن يختفي بسهولة. المشبك ذو القيمة
المستمرة 1.0 له نفس تأثير المشبك مع قيمة ثبات عتبة ، ولكن ليس من
السهل نسيانه. يسمح استخدام قيم الاستمرارية العددية بالتعلم عبر الإنترنت
في ظل وجود ضوضاء. قد يكون نمط الإدخال غير المرئي سابقًا عبارة عن ضوضاء
، أو قد يكون بداية اتجاه جديد يتكرر في المستقبل. من خلال تطوير نقاط
الاشتباك العصبي الجديدة ، يمكن للشبكة أن تبدأ في تعلم أنماط جديدة في
المرة الأولى التي تواجهها فيها ، ولكنها تأخذ سلوكيات مختلفة فقط بعد عدة
عروض تقديمية للنمط الجديد. زيادة المثابرة إلى ما بعد العتبة تعني أن تجربة
أنماط أكثر من الأنماط الأخرى ستستغرق وقتًا أطول في النسيان.
تعتمد الخلايا العصبية HTM وشبكات HTM على نمط توزيع نشاط الخلية ، لذا
فإن قوة تنشيط أي خلية عصبية أو مشابك ليست مهمة جدًا. لذلك ، في محاكاة
HTM ، نستخدم حالات ثنائية لمحاكاة تنشيط الخلايا العصبية والأوزان المشبكية.
بالإضافة إلى ذلك ، من المعروف أن المشابك البيولوجية عشوائية (فيصل
وآخرون ، 2008) ، لذا لا يمكن أن تتطلب نظرية القشرة المخية الحديثة فعالية
متشابكة دقيقة. على الرغم من أن الحالات والأوزان العددية قد تحسن الأداء ، إلا
أنها ليست ضرورية من وجهة نظر نظرية ، وجميع عمليات المحاكاة لدينا تعمل
بشكل جيد بدونها. يتم تقديم قواعد التعلم الرسمية المستخدمة في محاكاة
شبكة HTM الخاصة بنا في قسم المواد والأساليب.
نتائج المحاكاة
يوضح الشكل 6 أداء شبكة عصبون HTM التي تنفذ ذاكرة التسلسل عالية
الترتيب. تتكون الشبكة المستخدمة في الشكل 6 من 2048 عمودًا صغيرًا ،
يحتوي كل منها على 32 خلية عصبية. يحتوي كل خلية عصبية على 128 مقطعًا
شجيريًا قاعديًا ، ويحتوي كل جزء شجيري على ما يصل إلى 40 نقطة تشابك
عصبية فعلية. نظرًا لأن هذه المحاكاة تستخدم فقط لتوضيح خصائص ذاكرة
التسلسل ، فإنها لا تتضمن المشابك القمية. تعرض هذه الشبكة جميع السمات
الخمس المطلوبة لذاكرة التسلسل المدرجة سابقًا.
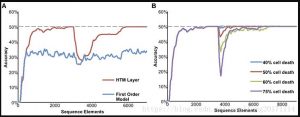
الشكل 6. نتائج محاكاة شبكة الذاكرة التسلسلية. يحتوي دفق الإدخال
المستخدم لهذا الرسم البياني على تسلسلات عالية الترتيب مختلطة بعناصر
عشوائية. الحد الأقصى لمتوسط دقة التنبؤ الممكنة لدفق البيانات هذا هو 50٪.
(أ) التعلم المتقدم عبر الإنترنت. يوضح الخط الأحمر أن الشبكة تعلمت وحققت
أقصى أداء ممكن بعد حوالي 2500 عنصر تسلسل. في العنصر 3000 ، يتغير
التسلسل في دفق البيانات. عندما يتعلم النموذج هيكلًا زمنيًا جديدًا ، تنخفض
دقة التنبؤ ثم تتعافى. للمقارنة ، يظهر الأداء الأقل لشبكة الترتيب الأول باللون
الأزرق. (ب) تدمر الشبكة المتانة. بعد أن وصلت الشبكة إلى أداء مستقر ، قمنا
بإلغاء تنشيط الخلايا العصبية المختارة عشوائيًا. عندما يصل معدل موت الخلايا
إلى 40٪ ، لا يوجد أي تأثير تقريبًا على الأداء. عندما يتجاوز معدل موت الخلية
40٪ ، ينخفض أداء الشبكة ، لكنها تتعافى عندما تعيد الشبكة استخدام الخلايا
العصبية المتبقية.
على الرغم من أننا قمنا بتطبيق شبكات HTM على العديد من أنواع البيانات
الحقيقية ، في الشكل 6 ، نستخدم مجموعات بيانات اصطناعية لتوضيح
خصائص الشبكة بشكل أوضح. الإدخال عبارة عن دفق من العناصر ، حيث يتم
تحويل كل عنصر إلى أعمدة صغيرة منخفضة النشاط بنسبة 2٪ (40 من إجمالي
2048 عمودًا نشطًا). تتعلم الشبكة نموذجًا تنبئيًا للبيانات بناءً على التحولات
الملحوظة في دفق الإدخال. في الشكل 6 ، يحتوي تدفق البيانات الذي يتم
تغذيته على الشبكة على مزيج من العناصر العشوائية والتسلسلات المتكررة.
يتكون التسلسل المضمن من ستة عناصر في الطول ويتطلب سياقًا زمنيًا عالي
المستوى لتحقيق توضيح كامل وأفضل دقة للتنبؤ ، مثل “XABCDE” و
“YABCFG”. بالنسبة لهذه المحاكاة ، نقوم بتصميم تدفق بيانات الإدخال بحيث
يكون الحد الأقصى لمتوسط دقة التنبؤ الممكنة هو 50٪ ، والذي لا يمكن تحقيقه
إلا باستخدام تمثيلات عالية الترتيب.
يوضح الشكل 6 أ التعلم عبر الإنترنت والتنبؤ المتقدم. تظهر دقة التنبؤ لشبكة
HTM بمرور الوقت باللون الأحمر. تبدأ دقة التنبؤ من الصفر وتزداد كلما وجدت
الشبكة أنماطًا زمنية متكررة مختلطة في انتقالات عشوائية. للمقارنة ، تظهر
دقة شبكة الترتيب الأول (التي تم إنشاؤها باستخدام خلية واحدة فقط لكل
عمود) باللون الأزرق. بعد التعلم الكافي ، حققت شبكة HTM عالية الترتيب 50٪
من أقصى دقة تنبؤ ممكنة ، بينما وصلت دقة الشبكة من الدرجة الأولى إلى
33٪ فقط. بعد أن تصل الشبكة إلى أقصى أداء لها ، يتم تعديل التسلسل
المضمن. في هذا الوقت ، تنخفض الدقة ، ولكن نظرًا لأن الشبكة تتعلم
باستمرار ، تتم استعادتها من خلال تعلم وضع ترتيب أعلى جديد.
يوضح الشكل 6B قوة الشبكة. بعد أن وصلت الشبكة إلى أداء مستقر ، قمنا
بإلغاء تنشيط الخلايا العصبية المختارة عشوائيًا. عندما يكون معدل موت الخلايا
مرتفعًا مثل حوالي 40٪ ، يكون التأثير على الأداء ضئيلًا. ترجع هذه القوة إلى
تحمل الضوضاء الموصوف سابقًا ، والذي يحدث عندما تتجاوز المشابك التي
تشكلها الأجزاء المتغصنة نقاط الاشتباك العصبي اللازمة لإنتاج ارتفاع NMDA.
عند المستويات الأعلى من موت الخلايا ، سينخفض أداء الشبكة مبدئيًا ، ولكن
نظرًا لأن الشبكة تستخدم الخلايا العصبية المتبقية لإعادة تعلم النمط ، فسوف
يتعافى أداء الشبكة.
كما هو موضح في جزء من الشكل 6 ب ، النموذج قوي جدًا وغير حساس تمامًا
لإعدادات المعلمات المختلفة. أهم المعلمات هي عتبة الذروة التغصنية وعدد
نقاط الاشتباك العصبي المخزنة في كل وضع. عند تعيين هذه المعلمات ، تأكد
من تذكر الجدول الموجود في نص S1 ، والذي يسرد المعلمات المتعلقة بمعدل
الخطأ المنخفض. من المهم بشكل خاص التأكد من أن العتبة التغصنية
والمشابك لكل وضع مرتفعان بدرجة كافية (على الأقل 12 و 20 على التوالي).
كما ذكرنا سابقًا ، ترتبط هذه الأرقام ارتباطًا وثيقًا بالأدبيات التجريبية. يمكن العثور
على مناقشة أكثر تفصيلاً لهذه المعادلات في Ahmad و Hawkins (2016).
تتضمن بعض المواد والطرق معلمات محددة مستخدمة في هذه التجارب.
مناقشة
نقترح نموذجًا للخلايا العصبية الهرمية ، والذي يختلف تمامًا عن نموذج العصبون
المستخدم في معظم الشبكات العصبية الاصطناعية. السمة الرئيسية للخلايا
العصبية النموذجية هي استخدامها للتشعبات النشطة ومناطق التكامل
المتشابك المتعددة (القريبة والقاعدية والقمية). تسمح التشعبات النشطة
للخلايا العصبية بالتعرف بشكل موثوق على مئات الأنماط المستقلة في
مجموعات الخلايا الكبيرة. تلعب منطقة التكامل المشبكي دورًا فريدًا في
الوظيفة ، مما يمكّن الخلايا العصبية من توقع التحولات والتسلسلات في نشاط
الخلية. في هذا النموذج ، تتسبب المشابك القريبة فقط بشكل مباشر في
إمكانات الفعل ، والأنماط التي تم اكتشافها على التشعبات القاعدية والقمية
تزيل استقطاب الخلية ، مما يمثل التنبؤات.
وجدنا أن شبكة هذه الخلايا العصبية المدمجة مع التثبيط المحلي السريع يمكنها
معرفة التسلسل في دفق البيانات. يتنبأ اكتشاف المشبك القاعدي بالنمط
السياقي للإدخال الأمامي التالي. يتنبأ الكشف عن المشابك القمي بنمط
التغذية الراجعة للتسلسل بأكمله. يعتمد تشغيل الخلايا العصبية والشبكات على
نشاط عصبي متناثر. يتعلم نموذج الذاكرة المتسلسلة باستمرار ، ويستخدم
سياق زمني متغير للتنبؤ ، ويمكنه إجراء تنبؤات متعددة في وقت واحد ،
ويستخدم قواعد التعلم المحلية فقط ، وهو قوي لعناصر الشبكة ، والضوضاء ،
وفشل تغييرات الوضع.
لقد تم اقتراح أن الخلايا العصبية ذات التشعبات النشطة يمكن تشكيلها بشكل
مكافئ باستخدام مدركات متعددة الطبقات (Poirazi et al. ، 2003). لذلك ، فإن
الفوائد الوظيفية والنظرية للتشعبات النشطة غير واضحة. يقترح نموذج ذاكرة
التسلسل الموضح في هذه المقالة هذه الفوائد من خلال تعيين أدوار فريدة
لمناطق تكامل متشابك مختلفة. على سبيل المثال ، لا يتم تنشيط الخلايا
العصبية الهرمية النموذجية الخاصة بنا إلا بشكل مباشر من خلال الأنماط
المكتشفة على المشابك القريبة ، بينما بالنسبة للأنماط المكتشفة على
التشعبات القاعدية والقمية ، فإنها تحافظ على استقطاب أكثر ديمومة للعتبة
الفرعية. علاوة على ذلك ، فإن التأثير المثبط للشبكة ينطبق بشكل مختلف على
مناطق التكامل المتشابك المختلفة. أخيرًا ، تختلف قواعد التعلم غير الخاضعة
للإشراف وتعمل على نطاقات زمنية مختلفة وفقًا لمنطقة التكامل. على الرغم
من أنه من الممكن تصور إنشاء دائرة من الخلايا العصبية الحسية المعيارية التي
تحتوي على كل هذه العمليات ، فإننا نقترح استخدام نموذج عصبون يحتوي على
تشعبات نشطة ومنطقة تكامل فريدة هو نهج أكثر أناقة وأضيق الحدود. كما أنه
يعكس البيولوجيا الأساسية بشكل أوثق.
العلاقة مع النموذج السابق
من المفيد مقارنة آلية ذاكرة التسلسل البيولوجي المقترحة مع تقنيات ذاكرة
التسلسل الأخرى المستخدمة في مجال التعلم الآلي. الأسلوب الأكثر شيوعًا
هو نماذج ماركوف المخفية (HMMs) (رابينر وخوان ، 1986). يستخدم HMM
على نطاق واسع ، خاصة في التعرف على الكلام. نموذج HMM الأساسي هو
نموذج من الدرجة الأولى ، ودقته مماثلة لنموذج الترتيب الأول الموضح في
الشكل 6 أ. يمكن لتغيير HMM نموذج التسلسل المقيد عالي الترتيب عن طريق
ترميز حالة الترتيب العالي يدويًا. تسمح الشبكات العصبية المتأخرة بالوقت
(TDNN) (Waibel ، 1989) للشبكات العصبية المغذية بمعالجة مجموعة فرعية
محدودة من التسلسلات عالية الترتيب من خلال الجمع الصريح بين المدخلات
المتأخرة. في الآونة الأخيرة ، أصبحت الشبكات العصبية المتكررة ، وخاصة
الذاكرة طويلة المدى (LSTM) (Hochreiter and Schmidhuber ، 1997) ، شائعة
وتتفوق بشكل عام على HMM و TDNN. على عكس شبكات HTM ، لا تحاول
HMM و TDNN و LSTM نمذجة الكائنات الحية بأي تفاصيل ؛ لذلك ، لديهم القليل
من الفهم لوظيفة الخلايا العصبية أو القشرة الحديثة. تتمثل المزايا الوظيفية
الرئيسية لنموذج HTM على هاتين التقنيتين في قدرته على التعلم المستمر
وقوته الممتازة وقدرته على عمل تنبؤات متعددة في وقت واحد. يمكن العثور
على مقارنة أكثر تفصيلاً في الجدول S1 و Cui et al. (2015).
هناك العديد من نماذج ذاكرة تسلسل التحفيز البيولوجي ذات الصلة. تمت
مناقشة اكتشاف المصادفة في الخلايا الهرمية في سياق ارتباط القشرة
المهادية الصدغية وتذبذبات 40 هرتز (Llinás et al. ، 1994). درست العديد من
الأوراق نماذج العصبونات المتصاعدة في سياق التعلم المتسلسل (ماس ،
1997 ؛ دينيف ، 2008 ؛ Ghosh-Dastidar and Adeli ، 2009 ؛ Jahnke et al. ،
2015). هذه النماذج هي فيزيائية حيوية أكثر من النماذج العصبية المستخدمة
في أدبيات التعلم الآلي. لقد أظهروا كيف تجعل اللدونة المعتمدة على الزمن
(STDP) الخلايا تستجيب لتسلسل محدد من الطفرات قبل المشبكية وتأخير
زمني محدد بين كل ارتفاع (روف وشميت ، 1997 ؛ راو وسيجنوفسكي ، 2000 ؛
جوتيج و Sompolinsky ، 2006). ميميشيمر وآخرون. أظهر (2014) أنه يمكن
تعلم العديد من التسلسلات المحددة التوقيت بدقة وإعادة تشغيلها ، والتي
يمكن استخدامها لنمذجة إخراج الصوت الغني للطيور المغردة. عادةً ما تقتصر
هذه النماذج على تسلسلات ماركوف (غير عالية الترتيب) ولا يتم تطبيقها على
مهام العالم الحقيقي المعقدة.
بشكل عام ، مستوى تفاصيل نموذج العصبونات المرتفعة أقل من نموذج HTM
المقترح في هذه المقالة. إنهم يصوغون بشكل صريح وقت تكامل إمكانات ما بعد
المشبكي ، والتأخير الزمني المقابل عادة ما يكون أقل من ملي ثانية إلى عدة
ميلي ثانية. عادةً ما يتعاملون أيضًا مع مجموعة فرعية صغيرة جدًا من نقاط
الاشتباك العصبي على الخلايا العصبية ولا يقومون بشكل صريح بنمذجة
التشعبات النشطة غير الخطية أو مناطق التكامل المتشابك المتعددة (ولكن
انظر Legendstein and Maass ، 2011). تركيز عملنا على نطاق أوسع. تحتوي
الخلايا العصبية لمحاكاة العمل الموصوفة في هذه المقالة على مجموعة كاملة
من نقاط الاشتباك العصبي والتشعبات النشطة ومناطق تكامل متشابك متعددة.
تحتوي الشبكة على عشرات الآلاف من الخلايا العصبية المرتبة في أعمدة
وطبقات. النموذج الناتج هو ذاكرة تسلسل معقدة حسابيًا ، والتي يمكن تطبيقها
على مشاكل العالم الحقيقي (Cui et al. ، 2015). أحد قيود نموذج HTM المقترح
في هذه المقالة هو أنه لا يتعامل مع التوقيت المحدد للتسلسل. لذلك ، فإن
الاتجاه المثير للاهتمام للبحث في المستقبل هو ربط هذين المستويين من
النمذجة ، أي إنشاء نماذج فيزيائية حيوية مفصلة تعمل على مستوى طبقة خلية
كاملة. أبلغ Billaudelle and Ahmad (2015) عن بعض التقدم ، ولكن لا يزال
هناك الكثير من العمل الذي يتعين القيام به في هذا المجال.
قدرة الشبكة والتعميم
كان هناك عمل مهم في الماضي في فهم قدرات الأنظمة ذات الخلايا العصبية
الخطية وغير الخطية والعناصر الحسابية (Cover، 1965؛ Vapnik et al.، 1994)
ومعدلات الخطأ المقابلة لها ( هوسلر ، 1988 ، 1990). تمت دراسة الجهاز
العصبي المتناثر في Kanerva (1988) ، Olshausen and Field (1997) ، وعمل
حديث مثل دراسة Jahnke et al. (2015). لم تتضمن الأدبيات حتى الآن توصيفًا
كاملاً لمعدل الخطأ مع تمثيلات متفرقة للمعلمات المقابلة للخلايا العصبية
القشرية. في هذه الورقة ، قمنا بتوسيع عملنا السابق لإظهار أنه يمكن تحديد
الأنماط المتفرقة عالية الأبعاد بشكل موثوق بينما تتطلب عددًا صغيرًا من نقاط
الاشتباك العصبي لبدء طفرات NMDA على التشعبات النشطة.
تمت دراسة إمكانيات الأشكال المختلفة لذاكرة التسلسل من قبل
(Sompolinsky and Kanter ، 1986 ؛ Riedel et al. ، 1988 ؛ Leibold and
Kempter ، 2006). في نموذجنا ، يتم الحصول على تقدير سعة التسلسل
مباشرة. على الرغم من أننا نشير إلى نموذج الشبكة على أنه “ذاكرة
متسلسلة” ، إلا أنها في الواقع ذاكرة محولة. لا يوجد تمثيل أو مفهوم لطول
التسلسل أو عدد التسلسلات المخزنة. تتعلم الشبكة فقط التحويل بين
المدخلات. لذلك ، يتم قياس سعة الشبكة من خلال عدد التحويلات التي يمكن
لشبكة معينة تخزينها. يمكن حساب ذلك على أنه نتاج دورة العمل المتوقعة
لخلية عصبية واحدة (الخلايا لكل عمود / تناثر الخلايا) مضروبًا في عدد الأنماط
التي يمكن لكل خلية عصبية التعرف عليها في التشعبات القاعدية. على سبيل
المثال ، 2٪ من الأعمدة في الشبكة نشطة ، ولكل عمود 32 خلية ، وتتعرف كل
خلية على 200 نمط على التشعبات الأساسية ، والتي يمكنها تخزين ما يقرب
من 320،000 تحويل ((32 / 0.02) * 200). السعة لها علاقة خطية مع عدد
الخلايا لكل عمود وعدد الأنماط التي تتعرف عليها المشابك القاعدية لكل خلية
عصبية.
يمكن أن يمثل نموذجنا تسلسلات معقدة عالية الترتيب (غير ماركوفية). يمكن
للنموذج أن يتعلم تلقائيًا الاعتماد على الوقت البعيد جدًا. لذلك ، فإن مقياس
السعة المهم هو عدد المرات التي يمكن أن يظهر فيها إدخال معين في سياقات
زمنية مختلفة دون التباس. هذا مشابه لعدد المرات التي تظهر فيها قطعة
موسيقية معينة في اللحن دون تشويش ، أو عدد المرات التي يمكن فيها تذكر
كلمة معينة في جمل مختلفة. إذا كان العمود المصغر يحتوي على 32 خلية ،
فهذا لا يعني أن وضعًا معينًا يمكن أن يحتوي فقط على 32 تمثيلًا مختلفًا. على
سبيل المثال ، إذا افترضنا أن كل إدخال يحتوي على 40 عمودًا نشطًا ، ولكل
عمود 32 خلية ، ولكل عمود خلية نشطة واحدة ، فإن كل نمط إدخال يحتوي
على 3240 تمثيلًا محتملاً ، وهو في الواقع عدد لا نهائي. لذلك ، فإن الحد
الفعلي ليس تمثيليًا ، ولكنه يعتمد على الذاكرة. تعتمد السعة على عدد
التحويلات التي يمكن تعلمها باستخدام مجموعة معينة من الأعمدة المتفرقة.
حتى الآن ، ناقشنا الطبقة الخلوية فقط ، حيث يمكن توصيل جميع الخلايا في
الشبكة بجميع الخلايا الأخرى ذات الاحتمال نفسه. هذا ينطبق على الشبكات
الصغيرة ، ولكن ليس على الشبكات الكبيرة. في القشرة المخية الحديثة ، من
المعروف أن معظم المناطق لديها تنظيم طوبولوجي. على سبيل المثال ، لا
تتلقى الخلايا الموجودة في المنطقة V1 سوى مدخلات تلقائية من جزء صغير
من شبكية العين ، ولا تتلقى سوى مدخلات جانبية من منطقة محلية من V1.
يمكن تكوين شبكة HTM بهذه الطريقة عن طريق ترتيب الأعمدة في صفيف
ثنائي الأبعاد واستخدام توزيع احتمالية ثنائية الأبعاد تتمحور حول الخلايا العصبية
لتحديد نقاط الاشتباك العصبي المحتملة لكل شجيري. يمكن أن تكون الشبكة
المنظمة طوبولوجيًا كبيرة بشكل تعسفي.
أحد الاعتبارات الرئيسية لخوارزميات التعلم هو مشكلة التعميم أو القدرة على
التعامل مع الأنماط الجديدة بقوة. يتم تعلم آلية ذاكرة التسلسل التي حددناها
من خلال تكوين عينة صغيرة من نقاط الاشتباك العصبي للخلايا العصبية النشطة
في تيار من الأنماط المتفرقة. تسمح خصائص التمثيل المتناثر بشكل طبيعي
بمثل هذه التعميمات المنهجية. سيكون هناك تداخل ضئيل للغاية في نمطين
متناثرين تم اختيارهما عشوائيًا. حتى التداخل الصغير (مثل 20٪) مهم جدًا
ويعني أن التمثيل له معنى دلالي مهم. العتبة التغصنية أقل من العدد الفعلي
لنقاط الاشتباك العصبي في كل جزء ، لذلك ستتعرف المقاطع على الأنماط
الجديدة المتشابهة ذات الصلة اللغوية. سيرى النظام أوجه التشابه بين
التسلسلات المختلفة ويقوم بعمل تنبؤات جديدة بناءً على القياس.
تنبؤ قابل للاختبار
تحتوي هذه النظرية على عدة تنبؤات قابلة للاختبار.
توفر هذه النظرية تفسيرًا حسابيًا للظاهرة التي تمت ملاحظتها تجريبيًا ، أي أن
نشاط الخلية الكلي يصبح قليلًا أثناء التدفق الحسي المستمر والمتوقع (Vinje
and Gallant، 2002؛ Yen et al.، 2007؛ مارتن وشرودر ، 2013). بالإضافة إلى
ذلك ، يتوقع أن يؤدي الإدخال غير المتوقع إلى زيادة نشاط الخلية ، والتي يجب
أن تكون مرتبطة رأسياً في الأعمدة المصغرة. من ناحية أخرى ، ستؤدي
المدخلات المتوقعة إلى أنشطة غير ذات صلة في الأعمدة المصغرة. تجدر
الإشارة إلى أن الحانات الصغيرة ليست مطلبًا صارمًا للنظرية. يتطلب هذا
النموذج فقط وجود مجموعات صغيرة من الخلايا التي تشترك في استجابة
تلقائية وتمنع بعضها البعض. نسمي هذه المجموعات أعمدة مصغرة ، لكن
الجانب العمودي ليس ضروريًا ، ويمكن أن يكون التجميع مستقلاً عن الأعمدة
المصغرة الفعلية.
التنبؤ الأساسي الثاني للنظرية هو أن النمط الحالي لنشاط الخلية يحتوي على
معلومات حول المنبهات السابقة. أبلغ نيكوليتش وزملاؤه عن نتائج تجريبية مبكرة
تدعم هذا التوقع. (عام 2009). هناك حاجة إلى مزيد من البحث للتحقق من
الطبيعة الدقيقة لنشاط الخلية الديناميكي ودور الخلفية الزمنية في التسلسلات
عالية الترتيب.
يجب وضع اللدونة المشبكية في مقاطع شجيرية يتم نزع استقطابها عن طريق
إدخال متشابك ، ثم تمرير إمكانات العمل العكسي بعد وقت قصير. تم الإبلاغ عن
هذا التأثير (Losonczy et al. ، 2008) ، على الرغم من أن هذه الظاهرة لم يتم
إثباتها على نطاق واسع.
يجب أن يكون هناك عدد قليل جدًا من التكوينات بين محور عصبي معين وقطاع
شجيري معين ، من الناحية المثالية ، يكون المشبك مثيرًا واحدًا فقط. إذا كانت
المحاور المثيرة تجلب العديد من نقاط الاشتباك العصبي على مقربة من شجيري
واحد ، فإن الخلايا قبل المشبكية ستؤدي إلى طفرات NMDA. يمكن التسامح مع
اثنين أو ثلاثة أو حتى أربعة نقاط تشابك من محور عصبي واحد إلى جزء شجيري
واحد ، ولكن إذا كانت المحاور تجعل بشكل روتيني مقطعًا شجيريًا واحدًا أكثر
تشابكًا ، فسوف يسبب أخطاء. يبدو أن التعلم Hebbian الخالص يشجع على
تكوين نقاط الاشتباك العصبي المتعددة. لمنع حدوث ذلك ، نتوقع وجود آلية تمنع
بشكل فعال تشكيل نقاط الاشتباك العصبي المتعددة بعد إنشائها. يمكن للمحاور
أن تشكل نقاط الاشتباك العصبي على أجزاء متغصنة مختلفة من نفس العصبون
دون التسبب في مشاكل ، لذلك نتوقع أن هذه الآلية ستكون مكانية في
المنطقة المحلية من الأجزاء التغصنية أو المحور المحوري.
عندما تولد خلية منزوعة الاستقطاب بواسطة ارتفاع NMDA لاحقًا إمكانية فعل
من خلال إدخال قريب ، فإنها تحتاج إلى قمع جميع الخلايا المثيرة الأخرى القريبة.
يجب أن يكون هذا قمعًا سريعًا ، وربما ذروة واحدة. الخلايا الكابتة للسلة التي
يتم تحفيزها بسرعة هي المصدر الأكثر احتمالا لهذا الكبت السريع (Hu et al. ،
2014).
تحتاج جميع الخلايا في الشريط الصغير إلى معرفة الاستجابة الموجزة الشائعة.
يتطلب هذا آلية لتشجيع جميع الوحدات في العمود المصغر على أن تصبح
نشطة في نفس الوقت أثناء تعلم وضع التغذية إلى الأمام. عندما تكون خلية
واحدة أو أكثر خالية من الاستقطاب بشكل طفيف ، يبدو أن مطلب الإثارة
المتبادلة هذا غير متوافق مع متطلبات التثبيط المتبادل السابق. ليس لدينا
اقتراحات محددة حول كيفية تلبية هذين المطلبين ، لكننا نتوقع آلية تكون فيها
الخلايا الموجودة في عمود في بعض الأحيان مثيرة بشكل متبادل ، وأحيانًا تكون
مثبطة بشكل متبادل.
من المعروف أن المناطق القشرية المختلفة والطبقات المختلفة لها تغييرات.
يمكن أن يكون للخلايا الموجودة في طبقات مختلفة أطوال شجيرية مختلفة
وعدد من نقاط الاشتباك العصبي (طومسون وبانيستر ، 2003). نتوقع أنه على
الرغم من هذه الاختلافات ، فإن جميع الخلايا العصبية المثيرة تتعلم التحولات أو
التسلسلات. الخلايا العصبية التي تحتوي على عدد أقل من الأجزاء المتغصنة
وعدد أقل من نقاط الاشتباك العصبي ستتعلم انتقالات أقل. نقترح أن التغيرات
في عدد التشعبات والمشابك ناتجة إلى حد كبير عن عدد الأنماط التي يمكن
العثور عليها في البيانات ، بدلاً من الاختلافات الوظيفية للخلايا العصبية نفسها.
الخلايا العصبية الهرمية شائعة جدًا في الحُصين. لذلك ، قد يكون جزء من نموذج
الخلايا العصبية والشبكة الخاص بنا قابلاً للتطبيق على الحُصين. ومع ذلك ، فإن
قرن آمون معروف بالتعلم السريع ، والذي لا يتوافق مع المشابك العصبية
الجديدة ، لأن المشابك البالغة قد تستغرق عدة ساعات لتتشكل (Knott et al. ،
2002 ؛ Trachtenberg et al. ، 2002 ؛ Niell et al. ، 2004 ؛ Holtmaat and Si
Voboda ، 2009). يمكن تحقيق التعلم السريع في نموذجنا ، فإذا لم يكن الأمر
يتعلق بتطوير نقاط الاشتباك العصبي الجديدة ، فستحتوي الخلايا على عدد كبير
من المشابك غير النشطة أو “الصامتة” (Kerchner and Nicoll، 2008). ثم تعلم
سريعًا عن طريق تحويل المشبك الصامت إلى مشابك عصبية نشطة. عيب هذه
الطريقة هو أن الخلية تحتاج إلى مزيد من نقاط الاشتباك العصبي ، وهو مكلف
من الناحية الأيضية. تحتوي الخلايا الهرمية في CA2 للحصين على عدة أضعاف
عدد الخلايا الهرمية المشبكية في القشرة المخية الحديثة (Megías et al. ،
2001). إذا كانت معظم هذه المشابك صامتة ، فيمكن إثبات أن المنطقة CA2
تنفذ أيضًا متغير ذاكرة التسلسل الذي اقترحناه.
المصدر














